By Dr. Sarah Reed, UC Berkeley.
(Dr. Reed recently received her PhD, based in part on mima mound studies she carried out in the Reserve and on nearby ranchlands. In this second of two blogs, Sarah discusses the work she did for her dissertation, testing the biological hypothesis of Mima mound origin in the mound and pool grassland environment in and around the UC Merced Vernal Pool Reserve.)
Mima mound chronosequence study
I was introduced to Mima mounds my first year of graduate school and quickly became fascinated by their patterns and the mystery of their origin. After my first trip to the Mima mounds, on the very young UC Merced campus (there were only 2 buildings at that time!), I was hooked and decided to devote my thesis to studying the enigmatic bumps.
I structured the study around an observation that Alfred Wallace11, the co-founder of the theory of evolution, made of the California Mima mounds: “Those near the foothills are the largest, and they gradually diminish in size as they extend out into the plain.” Based on this, I selected a suite of soils in the Merced area which stretch from the foothills of the Sierra Nevada to the level surfaces of the Central Valley and which range in age from a few thousand to a few million years. As a chronologic sequence of soils (a chronosequence), they systematically vary in degree of soil weathering, as a result of varying exposure to the elements. By designing the study around this age gradient, I hoped to begin to unravel the linkages between soil formation processes, mound morphology and animal behavior.
I hypothesized that as soils age (and as the mobile soil layer thickness decreases), environmental pressure on organisms living in the soil (aka gophers) increases, prompting a response via increasingly directional (and thus constructional) sediment transport. This should lead to an increase in mound height and density if gophers are responding to decreasing habitability.
Spatial analysis of mound morphology
Using LIDAR (light detection and ranging) data, I quantified Mima mound abundance, size, and spatial pattern as a function of soil age, for nearly a quarter million mounds. I found that mound height and density increase systematically with soil age (and with decreasing depth to restrictive layer), consistent with the predictions of the biologic model12.
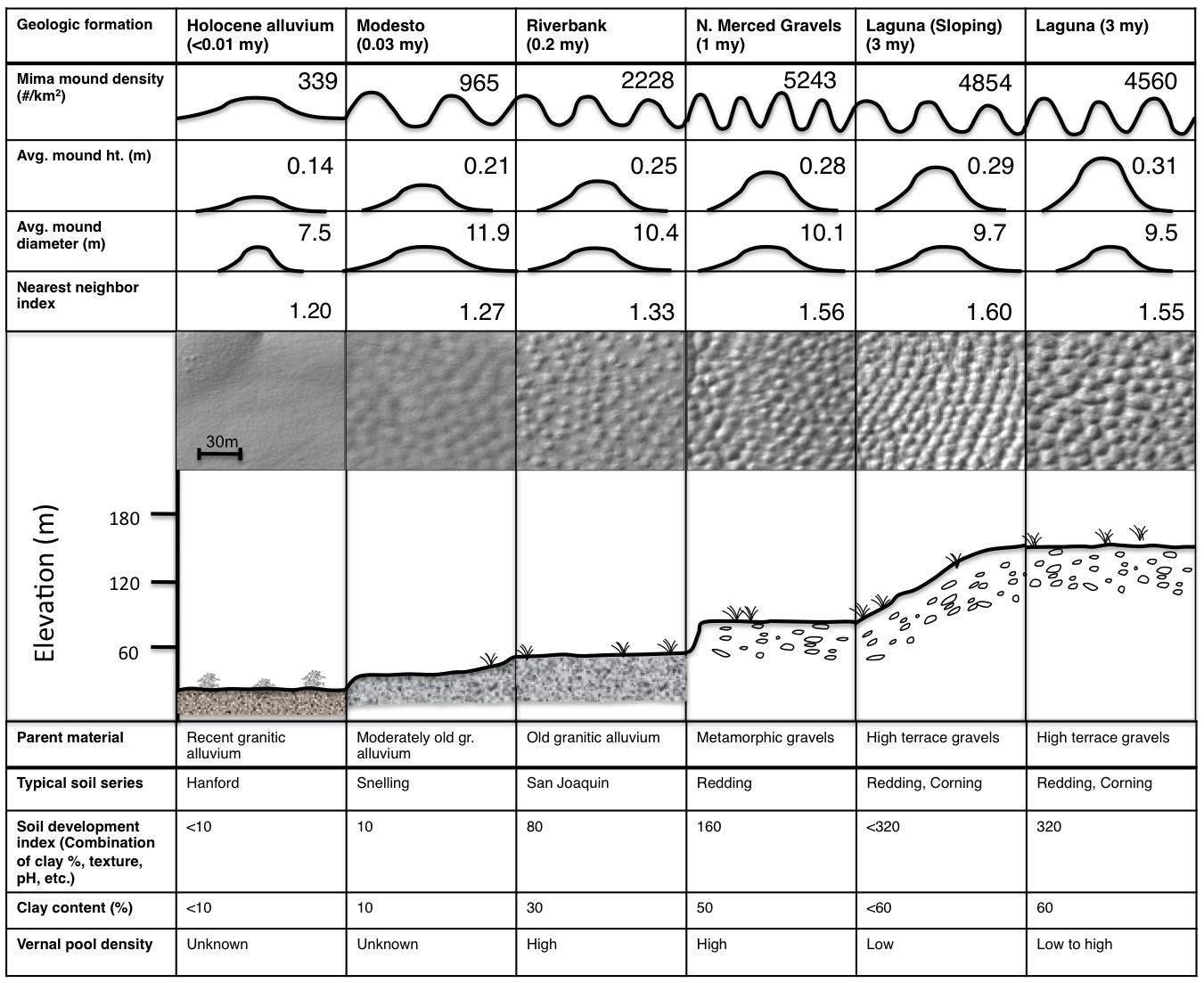
Table of LIDAR and environmental metrics across the Merced River soil chronosequence. Summary statistics in the upper portion of the table are based on data collected from 163775 mounds included in the chronosequence. Data in the bottom half of the table is from Marchand (1976), Harden (1987), and Vollmar (2002)13, 14 ,15. A larger analysis of 242343 mounds was conducted using soil depth to restrictive layer (usually hardpan) and the results also supported the biologic hypothesis (increasing mound height and density with decreasing soil depth).
In addition, I documented a pattern of strong regularity (as opposed to randomness or clustering) in mound distribution on all terraces of the Merced River chronosequence. Because gophers are highly territorial organisms, mounds should be uniformly spaced to reflect gopher home ranges (the area in which an animal lives and travels). The LIDAR data is consistent with this prediction. The average spacing between mounds (~12 m) 12 was quite similar to the spacing of gopher territories (~10 m) 16, and the average combined mound and swale areas in the Merced region (156 m2) is similar to the range of territory sizes for gophers in the San Joaquin Valley (120-250 m2) 17.
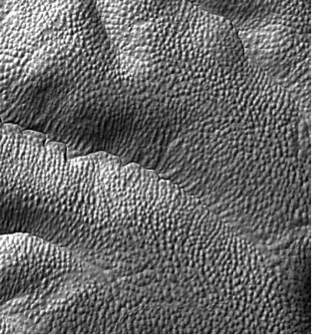
Figure 5. Shaded relief image showing the density of Mima mounds on the ~3 million year old Laguna terrace of the Merced River chronosequence, a few miles north of UCM campus. A single mound is approximately nine meters in diameter. North points to the top of the figure.
Gopher sediment transport
In addition to the LIDAR analysis, I also studied biotic sediment transport (how gophers move soil) by planting RFID (radio frequency identification) chips in Mima mound soil and quantifying the movement of the chips (and by proxy, the movement of soil) by gophers. Building on the work of Cox and Allen10, I documented direct evidence that gophers engage in constructional processes in Mima mound terrain and showed that as soils age, the net orientation of soil movement shifts upward and towards the center of mounds, as predicted by the biologic model.
Modeling the mounds
I also used two models to explore the biologic hypothesis of Mima mound origin. First, I used a hillslope model to estimate rates of Mima mound erosion and vernal pool deposition and compared the downwearing with rates of moundward-directed gopher sediment transport18. The results indicate that the rate of erosion is comparable to observed rates of sediment mounding via pocket gopher burrowing, suggesting that the mixing and churning of soil by living organisms likely plays a key role in maintaining Mima mound terrain. Second, I developed a model which approximates the energy required for the formation and maintenance of Mima mounds, and compared the amount to estimates of energy available to gopher populations in the Merced region19. The results show that the building of mounds by gophers is quite feasible (on both an energetic and a mass-based level) in relatively short time periods, strongly supporting a biotic origin for construction and preservation of Mima mounds and vernal pools.
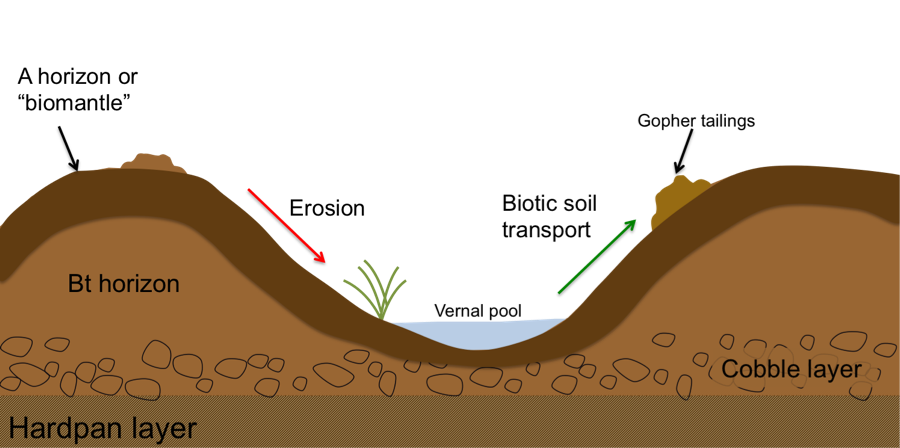
Figure 6. A conceptual model treating a Mima mound as a miniature hillslope, subject to erosion (red arrow), which occurs as a combination of physical and biological processes. For Mima mounds to exist and persist, there must be a restorative soil movement which counteracts the downslope erosion. The restorative movement is hypothesized to be due to upward (and moundward) sediment transport by gophers (green arrow).
Why does this matter to the vernal pools?
If the gopher hypothesis does prove correct, it leads to interesting questions about the interactions and feedbacks between landscape and life. That is, gophers respond, directly or indirectly, to very slow changes in soil development. Their response fundamentally changes the habitat in a way that facilitates the evolution of vernal pool plants and animals that are finely tuned to the small-scale topographic features (see, for example, Holland and Jain, 1984; Platenkamp, 1998; Bauder, 200520, 21, 22).
 Figure 7. Photo showing vernal pool in between several Mima mounds on the Laguna formation, northwest of UCM campus, on the Robinson Ranch.
Figure 7. Photo showing vernal pool in between several Mima mounds on the Laguna formation, northwest of UCM campus, on the Robinson Ranch. In these grassland habitats, the soil surface is likely mixed by burrowing animals, insects, and plant roots every several hundred years, with gravity directing the mixed material downslope. As such, the maintenance of strongly convex mounds with relatively soil-free, gravel-filled vernal pools points to the role of active restorative processes that maintain these landscapes. If gophers are indeed the creative and/or restorative agent, they must be considered as a possible keystone species of this ecosystem, and should be at the heart of long-term management strategies.
References
1 Johnson, D. L. and Horwath-Burnham, J. L. Introduction: Overview of concepts, definitions, and principles of soil mound studies. In Horwath-Burnham, J. and Johnson, D. L., editors, Mima Mounds: The Case for Polygenesis and Bioturbation, volume 490, pages 1–19. Geological Society of America, 2012.
2 Cox, G. W. The Distribution and Origin of Mima Mound Grasslands in San-Diego County, California. Ecology, 65(5):1397–1405, 1984a.
3 Washburn, A. L. Mima Mounds: An Evaluation of Proposed Origins with Special Refer- ence to the Puget Lowlands. Technical Report 29, Washington State Dept. of Natural Resources, Division of Geology and Earth Resources, 1988.
4 Higgins, C. Editorial comments on mounds. Geology, 18:284, 1990.
5 Dalquest, W. W. and Scheffer, V. B. The origin of the Mima mounds of western Washington. Journal of Geology, 50(1):68–84, 1942.
6 Grinnell, J. The burrowing rodents of California as agents in soil formation. Journal of Mammalogy, 4(3):137–149, 1923.
7 Nevo, E. Adaptive convergence and divergence of subterranean mammals. Annual Review of Ecology and Systematics, 10:269–308, 1979.
8 Patton, J. L. and Smith, M. F. Molecular evolution in Thomomys: phyletic systematics, paraphyly, and rates of evolution. Journal of Mammalogy, pages 493–500, 1981.
9 Lacey, E. A., Patton, J. L., and Cameron, G. N. Life underground: the biology of subterranean rodents. University of Chicago Press, 2000.
10 Cox, G. W. and Allen, D. W. Soil Translocation by Pocket Gophers in a Mima Moundfield. Oecologia, 72(2):207–210, 1987.
11 Wallace, A. R. The“Hog-Wallows”of California. Nature, 15:431–432, 1877.
12 Reed, S.E. (PhD Thesis) Pedologic-biologic feedbacks on the Merced River chronosequence: The role of pocket gophers (Thomomys bottae) in Mima mound-vernal pool ecosystems of the San Joaquin Valley. University of California, Berkeley / Pro-Quest, 2013. [Advisor: R.G. Amundson]
13 Marchand, D. Preliminary geologic maps showing Quaternary deposits of the Merced area, eastern San Joaquin Valley, 1976.
14 Harden, J. W. Soils developed in granitic alluvium near Merced, California. Geological Survey Bulletin (USA) 1590-A: Soil Chronosequences in the Western United States, pages A1–A65, 1987.
15 Vollmar, J. E. Landscape Setting . In Vollmar, J. E., editor, Wildlife and Rare Plant Ecology of Eastern Merced County’s Vernal Pool Grasslands, chapter 2, pages 7–54. University of California at Merced, 2002.
16Zinnel, K. C. and Tester, J. R. Plains pocket gopher social behavior. In Proceedings of the Thirteenth North American Prairie Conference. Windsor, Ontario, Canada, page 95ˆa“101, 1994.
17 Howard, W. E. and Childs, H. E. Ecology of pocket gophers with emphasis on Thomomys bottae mewa. University of California, California Agricultural Experiment Station, 1959.
18 Reed, S. E. and Amundson, R. G. Sediment, Gophers, and Time: A Model for the Origin and Persistence of Mima Mound-Vernal Pool Topography in the Great Central Valley. In Schlising, R. A. and Alexander, D. G., editors, Vernal Pool Landscapes, number 14 in Studies from the Herbarium, pages 15–27. California State University-Chico, 2007.
19 Reed, S. and Amundson, R. Using LIDAR to model Mima mound evolution and regional energy balances in the Great Central Valley, CA. In Horwath Burnham, J. and Johnson, D. L., editors, Mima Mounds: The Case for Polygenesis and Bioturbation, volume 490, chapter 1, pages 21–41. Geological Society of America, 2012.
20 Holland, R. F., and S. K. Jain. "Spatial and temporal variation in plant species diversity of vernal pools." Vernal pools and intermittent streams 28 (1984): 198-209.
21 Platenkamp, G. A. J. Patterns of vernal pool biodiversity at Beale Air Force Base. In Witham, C., Bauder, E., Belk, D., Ferren Jr, W., and Orndu↵, R., editors, Ecology, Con- servation, and Management of Vernal Pool Ecosystems-Proceedings froma 1996 conference. California Native Plant Society, Proceedings from a 1996 Conference, pages 151–160. Cal- ifornia Native Plant Society, 1998.
22 Bauder, E. T. The effects of an unpredictable precipitation regime on vernal pool hydrology. Freshwater Biology, 50(12):2129–2135, December 2005.
Date:
April 17, 2015

